
Mouse Fur Color
 The beach mouse, (Peromyscus polionotus) is a small rodent found in southeastern United States. It lives primarily in old fields and on white sand beaches. There are several sub-species, each having a somewhat unique fur color ranging from dark brown to very light brown, approaching white. The geography of fur colors is not random. The darkest colored subspecies tend to live inland while the lighter subspecies live on light sand beaches. This case will examine the evolution of light fur in beach mice from the molecular level up to the population genetics level.
The beach mouse, (Peromyscus polionotus) is a small rodent found in southeastern United States. It lives primarily in old fields and on white sand beaches. There are several sub-species, each having a somewhat unique fur color ranging from dark brown to very light brown, approaching white. The geography of fur colors is not random. The darkest colored subspecies tend to live inland while the lighter subspecies live on light sand beaches. This case will examine the evolution of light fur in beach mice from the molecular level up to the population genetics level.
Ecology: Predation and Camouflage
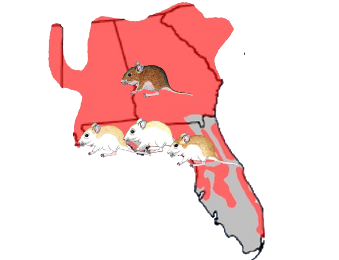 The beach mouse, (Peromyscus polionotus) is a small rodent found in southeastern United States. Different sub-species of beach mouse tend to live on different beaches in Florida’s panhandle region. One of the major differences among these subspecies is that they have different fur colors ranging from light to very dark.
The beach mouse, (Peromyscus polionotus) is a small rodent found in southeastern United States. Different sub-species of beach mouse tend to live on different beaches in Florida’s panhandle region. One of the major differences among these subspecies is that they have different fur colors ranging from light to very dark.
The distribution of fur colors is not random. The darkest colored subspecies tends to live inland where soils are dark brown while the lighter subspecies live on beaches where the sand is light brown.
Soil Color and Fur color
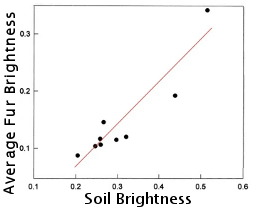
Ancestral populations of beach mouse likely had dark fur coats indicating that light fur color is a recent evolutionary event. Researchers have observed a very strong correlation between soil brightness and coat color among different populations (r = 0.92 p = 0.0004). This could indicate that there may be some kind of selection pressure that initially established, and currently maintains the differing coat colors among populations. Although the researchers who uncovered the relationship between fur color and soil brightness are skeptical about predation-related selection pressure (read more about their research here: Belk and Smith, 1996), other research indicates this may be occurring.
Selective Predation and Fur Color
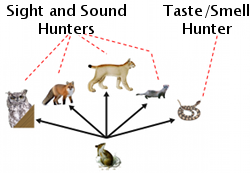
Most animals that hunt beach mice are sight and sound predators. When the mice are visually hunted, blending into the substrate can provide a significant advantage to help them avoid detection. When mice avoid predation they have a better opportunity to find a mate and pass their fur color alleles along to their offspring. Predation could thus be a driving force that changes fur color allele frequencies within mice populations to match soil or sand color.
To test this hypothesis, Kaufman (1974) set up two enclosures, one with dark soil and one with light. In each he tested the hunting behaviors of barn owls and screech owls. When both light and dark mice were released into enclosures, owls tended to catch the dark mouse first in light soil enclosures and the light mouse first in dark soil enclosures.


In a more recent study Vignieri et al (2010) made realistic clay models of light and dark mice and set them out in light and dark soil habitats to be attacked by predators. They found that light colored clay mice were frequently attacked in dark soil environments and dark colored clay mice were frequently attacked in light soil environments. Light colored clay mice in light soil environments and dark colored clay mice in dark soil environments were comparatively unharmed. (Picture taken from the Vignieri et al 2010; study found at http://www.ncbi.nlm.nih.gov/pubmed/20163447)
Cell Biology
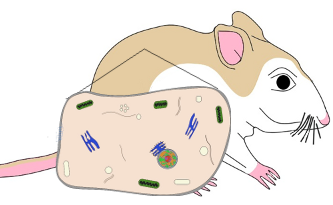 Different sub-species of the beach mouse (Peromyscus polionotus) have different fur colors. These differences are best described by examining melanocyte cell function. Melanocytes are located at the base of the hair papilla and make a pigment molecule called eumelanin. Pigment is then transported from the base to the hair cortex where coloration becomes visible.
Different sub-species of the beach mouse (Peromyscus polionotus) have different fur colors. These differences are best described by examining melanocyte cell function. Melanocytes are located at the base of the hair papilla and make a pigment molecule called eumelanin. Pigment is then transported from the base to the hair cortex where coloration becomes visible.
The Eumelanin Pigment
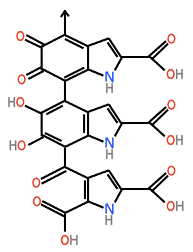 Eumelanin is a polymer of cross-linked dihydroxyindole carboxylic acids. It is composed of many units that can vary slightly in their composition.
Eumelanin is a polymer of cross-linked dihydroxyindole carboxylic acids. It is composed of many units that can vary slightly in their composition.
Different sub-species of the beach mouse (Peromyscus polionotus) have different fur colors. These differences are best described by examining melanocyte cell function. Melanocytes are located at the base of the hair papilla and make a pigment molecule called eumelanin. Pigment is then transported from the base to the hair cortex where coloration becomes visible.
Eumelanin Production
Activation of the MC1R protein occurs when it successfully binds to a molecule called an α melanocyte stimulating hormone (α-MSH). Alternatively, if the MC1R protein binds to a different molecule, an agouti stimulating peptide, the biosynthetic pathway leading to eumelanin synthesis may not occur. However, assuming that α-MSH binding occurs, cAMP synthesis begins. cAMP is a common gene regulator and it aids in the expression of enzymes necessary in the biosynthetic pathway leading to eumelanin – it stimulates the expression of four genes: c(Tyr), Tryp1, Tyrp2 and p. The c(Tyr) gene is responsible for an enzyme called tyrosinase that catalyzes a reaction in a biosynthetic pathway that converts the amino acid tyrosine into a molecule called dopaquinone. The Tyrp1 and Tryp2 genes hold the blueprint for tyrosine-related enzymes that catalyze reactions in a subsequent pathway that converts dopachrome into eumelanin. The p gene is responsible for a protein that also aids in this process. Eumelanosomes are then made in the golgi apparatus of the melanocyte that are then transported out of the melanocyte to the hair cortex.
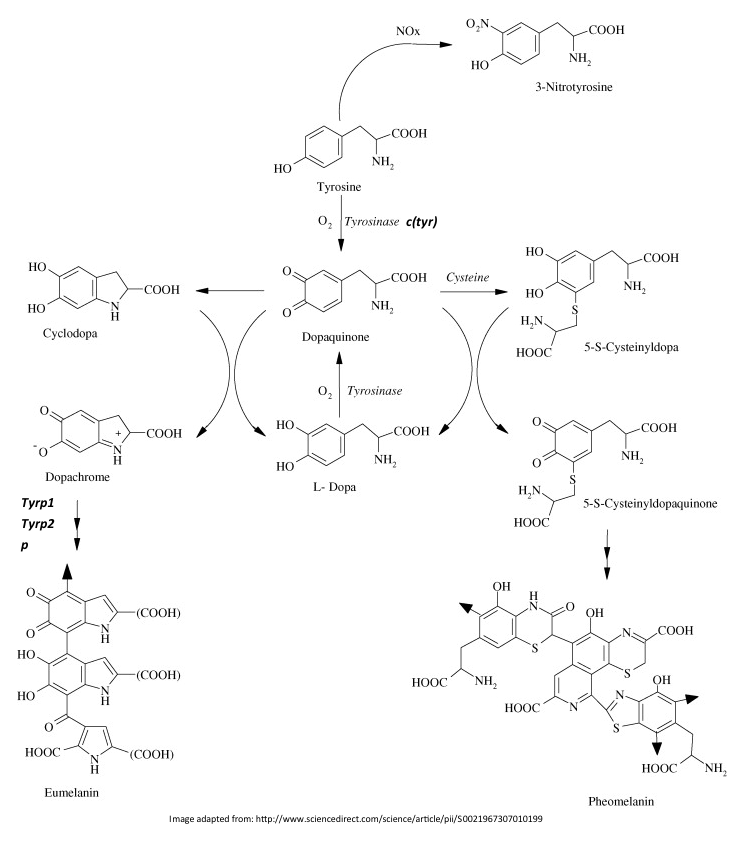
Without the successful binding of an α-MSH molecule by the MC1R protein, cAMP is not adequately regulated and the multistep biosynthetic pathway for eumelanin synthesis cannot take place. Structural changes to the MC1R protein as a result of a genetic mutation make it essentially non-functional and result in a different coat color due to changes in pigment production. This is often the case in coastal populations of beach mice where coat colors are lighter than their inland neighbors.
Molecular Genetics
Fur color in beach mice (Peromyscus polionotus) can be altered if the transmembrane protein, MC1R, has a mutation that disrupts its role in the synthesis of the eumelanin pigment. A functional MC1R protein binds to an α-melanocyte stimulating hormone that stimulates intracellular pigment production whereas a non-functional MC1R protein does not.
DNA, Central Dogma and the MC1R Protein
DNA is made up of long strings of nucleotides. There are four nucleotides in DNA: Adenine (A), Thymine (T), Guanine (G) and Cytosine (C). These nucleotides are held together by a phosphate-sugar “backbone” and are complemented by a parallel strand of nucleotides making a DNA molecule “double-stranded”. Parallel strands are held together by hydrogen bonds between bases; Adenine binds to Thymine, Cytosine binds to Guanine. Discrete sections of DNA carry sequences of nucleotides that, when processed, are responsible for proteins or functional RNA chains. These sections are called genes. When making a protein, the DNA is first transcribed into a single-stranded piece of messenger RNA (mRNA) using an enzyme called RNA polymerase. The newly made mRNA is then translated through a ribosome into a long chain of amino acids. This chain of amino acids folds up to make a functional protein. Proteins build the organism, including all measurable and observable characteristics.
The sequence of nucleotides that codes for the mRNA that produces the MC1R protein is shown below.

The Mutation
A single nucleotide substitution at position # 199 (cytosine to thymine) in the MC1R gene results in a non-functional MC1R protein. This small difference causes a change in the amino acid chain produced by translation. Amino acid at position #67 within the polypeptide chain becomes a cysteine rather than an arginine. This single change within the protein results in a conformation change that does not allow the MC1R protein to bind extracellular melanocyte stimulating hormone. When this binding does not occur eumelanin is not effectively produced within melanocyte cells (see Cell Biology for more details).
Population Genetics

In biological research, to say that something at the molecular or genetic level is responsible, a phenotypic trait, a plausible mechanism and an observable relationship should be identified. In the cell biology and molecular genetics sections we discuss a possible mechanism.
Gene and alleles
There are two different alleles that can be identified for the MC1R gene that aid in eumelanin production in beach mice (Peromyscus polionotus). An allele is a variant of a gene, which is a discrete section of DNA that codes for a polypeptide (i.e. protein) or a functional RNA molecule. In beach mice the MC1R gene has two alleles: R and C. The R allele results in a functional MC1R protein that stimulates eumelanin production. This polypeptide has amino acid arginine at position #67 due to the codon CGC at positions 199, 200 and 201 within the MC1R nucleotide sequence. The C allele results in a non-functional MC1R protein that does not stimulate eumelanin production. It has the amino acid cysteine at position #67 due to the codon TGC at positions 199, 200 and 201 within the MC1R nucleotide sequence.
Genotypes and Phenotypes
The MC1R gene is located on chromosome #16 in mice. Since every mouse cell has two copies of each autosomal chromosome, the cells of any given beach mouse will possess two copies of the MC1R gene – one on each chromosome. Since there are two MC1R alleles there are three possible genotypes that an individual may possess: RR, RC, and CC.
Correlating Coat color to Gene mutations
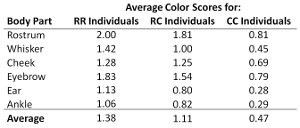
Hoekstra et al (2006) analyzed more than 400 mice to determine how fur color genotypes were related to fur color phenotypes. Mice were scored on a 2-point scale: 2 for very dark fur, 1 for medium, and 0 for very light. Scoring was done for seven different body parts – the whisker, the rostrum, the cheek, the eyebrow, the ear, the ventrum and the ankle. The scores were then matched to the known genotype that had been sequenced for each mouse, RR, RC, or CC.
They found that the RR mice were typically darker than the RC mice that in turn were typically darker than the CC mice.
Conclusion
Different alleles of the MC1R gene are responsible for some of the coat color differences observed among beach mouse populations. However, these alleles cannot explain all the color variation among populations and it is therefore probable that mutations in other genes and their alleles contribute to differences in coat color.