
Human Skin Pigmentation
 Human skin has many functions, one of which is mediating the effects of sunlight. Pigmentation, (skin color), in humans has evolved under selection pressure from the duration and intensity of sunlight. Ancestral populations near the equator were selected for dark skin, while those living in higher northern latitudes were selected for lighter skin. The distribution of “light” alleles of well-studied genes associated with skin color is non-random. Some of these “light” versions are found in most northern latitude populations; others are more geographically specific.
Human skin has many functions, one of which is mediating the effects of sunlight. Pigmentation, (skin color), in humans has evolved under selection pressure from the duration and intensity of sunlight. Ancestral populations near the equator were selected for dark skin, while those living in higher northern latitudes were selected for lighter skin. The distribution of “light” alleles of well-studied genes associated with skin color is non-random. Some of these “light” versions are found in most northern latitude populations; others are more geographically specific.
Most of the known genes (and alleles) for skin color either 1) are part of the biosynthetic pathway for melanin, or 2) regulate melanin biosynthesis. Complicating the story is polygenic inheritance. The expression of the many genes that contribute to skin color results in a continuum of skin color. This continuum becomes more pronounced as populations migrate and interbreed. There are no measurable genetic differences among and between human populations. Thus, the notion of “race” is a societal idea, not a biological trait.
Specialized Cells and their Functions
Both gene expression and natural selection are required for “building” traits of an organism, including skin color in humans. The outer cells of the skin provide protection from the sun by synthesizing and distributing pigment. Important in this process are melanocyte and keratinocyte cells.
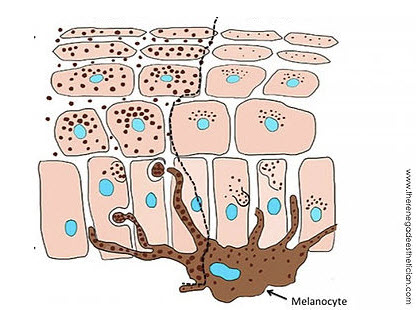
Figure (above): Melanocytes (brown) make and transport melanosomes for several “client” keratinocytes. They have long skinny “arms” to accomplish this. The Keratinocytes (pink) have variable numbers and sizes of melanosomes (black) that cluster around the nucleus.
Keratinocytes and Melanocytes
Human skin pigmentation is the result of gene expression and natural selection. Skin color requires the expression of dozens of genes all interacting to produce the characteristic of “skin color”. Differential reproductive success, the result of natural selection, ensures that an organism’s genes will be carried forward into grandchildren, great-grandchildren, great-great-grandchildren, etc.
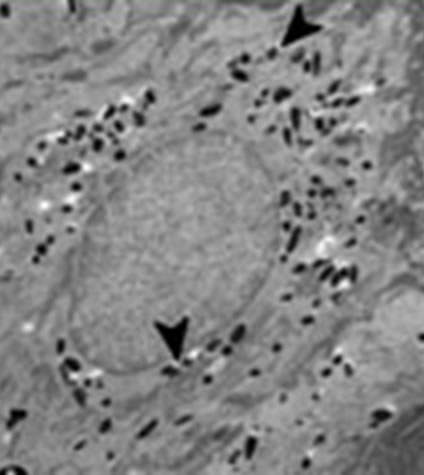
Figure (above): This electron microscope image shows melanosomes clustering near the nucleus of keratinocytes. They protect it from damaging ultraviolet rays by absorbing this relatively high energy light. Dark skin has more and larger melanosomes around the nucleus of keratinocytes that does light skin.
Skin has four functions: 1) act as a barrier between inside and outside worlds; 2) is a large sensory organ; 3) provides a home to an individual’s microbiome; and 4) interacts with the sun’s UV radiation. The outer layer of skin, the epidermis, is where skin color is expressed. The epidermis has two kinds of cells. Keratinocytes make up the outer layer of the skin and contain packages of pigment that are responsible for skin color. Melanocytes make the packages of pigment, melanosomes, and transfer them to the keratinocytes. There, they cluster around the nucleus. The number of melanosomes is related to the pigmentation or darkness of the skin. Melanosomes are transferred from melanocytes by either phagocytosis or endocytosis. Keratinocytes send paracrine signals to melanocytes to upregulate the formation of melanosomes.
Polygenic Inheritance
 Complicating the picture of gene expression is the many interacting genes/gene products involved in skin pigmentation. A simple model explains the influence that multiple genes can have on a characteristic. Complexity is compounded by other gene properties, such as multiple alleles and epistasis.
Complicating the picture of gene expression is the many interacting genes/gene products involved in skin pigmentation. A simple model explains the influence that multiple genes can have on a characteristic. Complexity is compounded by other gene properties, such as multiple alleles and epistasis.
An Analogy of Doses
 Skin pigmentation is the result of the interaction of many expressed genes: skin color is a polygenic characteristic. What is the nature of polygenic inheritance? This module uses a simple three-gene model, each with two alleles. The dominant form codes for a “dose” of dark skin and the recessive form codes for a “dose” of light skin. The darkest skin is due to six dominant “doses” and the lightest skin is due to six recessive “doses”. Varying combinations of the alleles result in seven discrete colors. Mendel’s laws (reviewed in slides) show that there are different combinations of the three expressed genes for each of the seven colors. The model shows the result of expressed alleles adding to a phenotype of one of seven discrete colors. However, skin color varies continuously in modern populations, a reflection of the many genes involved. Sorting out the role that each interacting gene product plays in the expression of skin color is a task not easily accomplished. Research has determined the role of a handful of genes, but not necessarily how these interact and certainly not how the constellation of genes involved in determining skin color interact. The problem is to determine the genetics of these tangled relationships.
Skin pigmentation is the result of the interaction of many expressed genes: skin color is a polygenic characteristic. What is the nature of polygenic inheritance? This module uses a simple three-gene model, each with two alleles. The dominant form codes for a “dose” of dark skin and the recessive form codes for a “dose” of light skin. The darkest skin is due to six dominant “doses” and the lightest skin is due to six recessive “doses”. Varying combinations of the alleles result in seven discrete colors. Mendel’s laws (reviewed in slides) show that there are different combinations of the three expressed genes for each of the seven colors. The model shows the result of expressed alleles adding to a phenotype of one of seven discrete colors. However, skin color varies continuously in modern populations, a reflection of the many genes involved. Sorting out the role that each interacting gene product plays in the expression of skin color is a task not easily accomplished. Research has determined the role of a handful of genes, but not necessarily how these interact and certainly not how the constellation of genes involved in determining skin color interact. The problem is to determine the genetics of these tangled relationships.

Figure (above): The pipe cleaners represent a duplicated pair of matched (homologous) chromosomes of a parent with what we’ll call the Ee genotype, ready to separate during meiosis I. The beads represent the alleles E (pink) and e (white). Upon separation the two kinds of alleles will segregate. During meiosis II, the duplicated alleles split apart.
The fact that one gene can have more than two alternative versions, or multiple alleles, also adds complexity. This is certainly the case for some of the known genes involved in skin color, such as MC1R and TRYP2. Additionally, the relationship between alleles is not always “dominant” and “recessive”. Rather, there is dominance variation, the most common of which is co-dominance. Epistasis, where modifier genes influence the expression of other genes, also contributes to the task of sorting out polygenic inheritance. For example, the genes responsible for determining the internal environment of melanosomes, where melanin pigment are made, regulate, or modify, the synthesis of melanin.
Genes and Proteins
Some genes expressed in the production of melanin are associated with 1) biosynthesis or 2) regulation of biosynthesis. Different alleles for these genes affect either the kind or amount of melanin.
Many Genes, Many Proteins
The expression of a multitude of genes is required for the characteristic known as skin color. This expression results in a continuous palette of skin color. The skin pigments, eumelanin and pheomelanin are synthesized in melanosomes, which form in melanocytes. Melanosomes subsequently are transported to keratinocytes where they protect those cells. The most completely studied genes related to human skin pigmentation are key to pigment synthesis and regulation of that synthesis.
Initiation of melanin synthesis: MC1R and ASIP
The hormone alpha-melanocyte stimulating hormone (α-MSH) interacts with the melanocortin 1 receptor, MC1R, on the outside membrane of melanocytes. cAMP levels increase. A cascade of reactions converts the amino acid tyrosine to the pigment eumelanin.
A single change in an amino acid lowers MC1R’s affinity for α-MSH. The result is the synthesis of the pigment pheomelanin instead of eumelanin and lighter skin. There are many alleles of the MC1R gene. Agouti Signaling Protein (ASIP, coded by the ASIP gene) is an antagonist to MC1R, shutting down the pathway of eumelanin synthesis. Thus, the production of pheomelanin is favored. Variations of this gene, alleles, are found in some people with lighter skin.
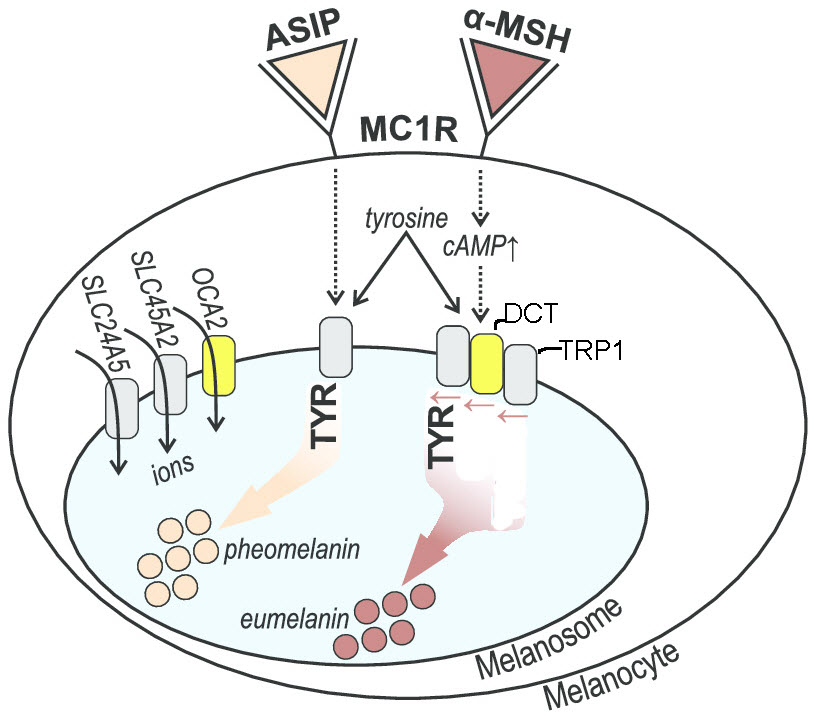
Synthesis of melanins: TYR, TRP1 and DCT (TRP2).
Tyrosinase, (coded by the TYR gene) is the primary enzyme involved in the conversion of tyrosine to melanin. Nucleotide differences in the TYR gene correlate with skin pigmentation variation in humans. There are two tyrosine related proteins that that work with tyrosinase to produce melanins. One is TRP1; the other is dopachrome tautomerase , DCT, which is also known as TRP2. These proteins are associated with regulating pigment synthesis, although the mechanisms are unclear. There are known alternative alleles for the corresponding genes, which appear to correlate with the kind and amount of melanin made.
Regulation of melanin synthesis: OCA2, SLC24A5, and SLC45A2 (=MATP)
The internal environment of the melanosome regulates the amount of melanin synthesized. Thus, genes and proteins that influence this environment are key in determining skin color. The SLC24A5 gene codes for a potassium dependent (K+) sodium/calcium (Na+/Ca++) exchange transporter. The amount of calcium in the melanosome affects the entry of tyrosine, which determines the amount of melanin made. A specific allele of this gene is common in light skin populations (Western Europe).
SLC45A2 (=MATP) regulates the melanosomal pH. pH levels affect tyrosinase activity and subsequently the amount of eumelanin or pheomelanin synthesized. There are many variations (alleles) in the SLC45A2 gene.
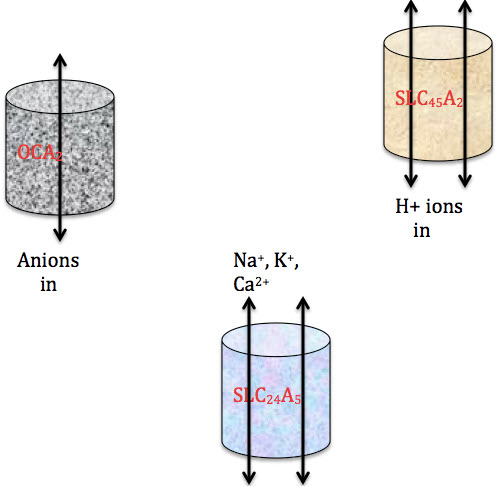 OCA2 (gene is OCA2) resembles anion transporters in bacteria and also regulates melanosomal pH. OCA2 is thought to serve as a control point at which ethnic skin color variation is determined. Thus, the internal environment determines the amount of either pigment made, regulated by transporters (p-protein, MATP, SLC24A5
OCA2 (gene is OCA2) resembles anion transporters in bacteria and also regulates melanosomal pH. OCA2 is thought to serve as a control point at which ethnic skin color variation is determined. Thus, the internal environment determines the amount of either pigment made, regulated by transporters (p-protein, MATP, SLC24A5
Selection
 The intensity of UV light was a strong environmental factor in the evolution of skin color. Dark indigenous people living near the equator had more offspring than their lighter counterparts; darker skin protects cell nuclei from damage. At higher latitudes, where UV light was limited, those populations with lighter skin and having the ability to synthesize vitamin D were favored.
The intensity of UV light was a strong environmental factor in the evolution of skin color. Dark indigenous people living near the equator had more offspring than their lighter counterparts; darker skin protects cell nuclei from damage. At higher latitudes, where UV light was limited, those populations with lighter skin and having the ability to synthesize vitamin D were favored.
Different Selection at Different Latitudes
There is a clear relationship between skin color and geography. Generally speaking, the darkest native people live in sub-Saharan Africa and in Melanesia. The lightest native people live in high northern latitudes. This pattern follows that of UV radiation, with the highest intensity radiation at or near the equator and the lowest at high latitudes. The ancestors of Homo sapiens probably had light skin. A major challenge to these populations in the African savannah was the sun. Those populations acquiring traits that helped them survive in their environment left more offspring than those that did not. Over time, Homo sapiens individuals in populations in Africa with darker skin (having lots of the pigment eumelanin) left behind more offspring than those with lighter skin. The three primary hypotheses for positive selection of dark skin are not mutually exclusive and relate to protecting DNA from UV damage in skin cell nuclei. These are 1) protecting skin from developing cancer; 2) preventing the photolysis of folate (vitamin B9); and 3) preventing skin, sweat glands and blood vessels from severe damage.
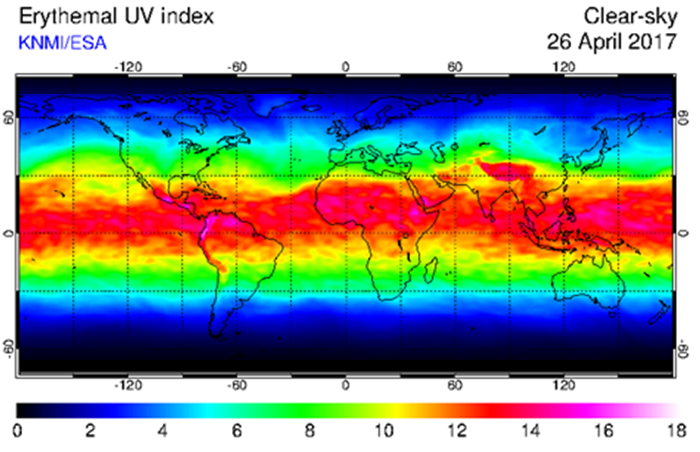
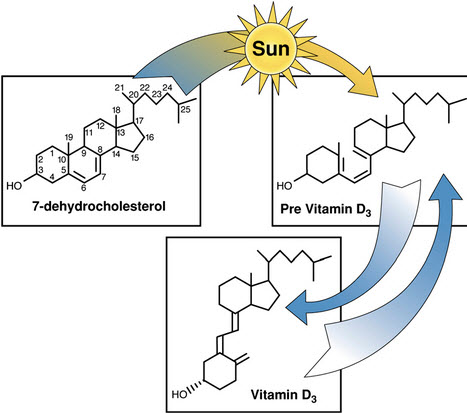 As humans migrated northward, the selection pressures for dark skin relaxed. Additionally, these populations met a new challenge: insufficient synthesis of Vitamin D, which requires the interaction of skin cells and UV light. Individuals with lighter skin could synthesize more vitamin D under low UV light conditions than those with darker skin. Their children were stronger than those that could not synthesize sufficient vitamin D. Modern societies fortify their food with both vitamin D and folate. This practice reflects the diversity of skin color in present day societies.
As humans migrated northward, the selection pressures for dark skin relaxed. Additionally, these populations met a new challenge: insufficient synthesis of Vitamin D, which requires the interaction of skin cells and UV light. Individuals with lighter skin could synthesize more vitamin D under low UV light conditions than those with darker skin. Their children were stronger than those that could not synthesize sufficient vitamin D. Modern societies fortify their food with both vitamin D and folate. This practice reflects the diversity of skin color in present day societies.
Population Genetics
 Within and between population genetic differences are used to characterize the distribution of variants in five genes associated with skin color across global populations. This information helps us understand the evolution of skin color in humans.
Within and between population genetic differences are used to characterize the distribution of variants in five genes associated with skin color across global populations. This information helps us understand the evolution of skin color in humans.
Genes and Alleles
 SNP (single nucleotide polymorphisms) and STR (short tandem repeats) data can be used to determine evolutionary relatedness among populations. These patterns of changes in DNA are the basis for the FST (fixation) statistic, an estimation of differences in populations based on genetic variation (π):
SNP (single nucleotide polymorphisms) and STR (short tandem repeats) data can be used to determine evolutionary relatedness among populations. These patterns of changes in DNA are the basis for the FST (fixation) statistic, an estimation of differences in populations based on genetic variation (π):
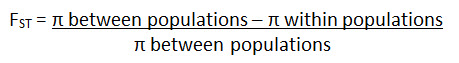
An FST value of 0 indicates that two populations are interbreeding; there is insignificant variation in the alleles of a gene under study. No evolution is occurring.
An FST value of 1 indicates that two populations are very different from one another as evidenced from the alleles of a gene under study. Evolution is probably occurring.
FST scores can be calculated for genes known to be involved in skin color pigmentation. These scores indicate how conserved or different these genes and their alleles are among and between populations.
Norton et. al (2007) studied the global distribution (a total of 53 populations, of which 27 are represented here) of variations of six genes associated with skin color: MC1R, ASIP, TYR, OCA2, MATP aka SLC45A2, and SLC24A5 . They found that the melanocortin 1 receptor (MC1R) variation was greater within populations than between populations: the FST value was small. Thus, the remainder of their analysis was based on the other five genes.
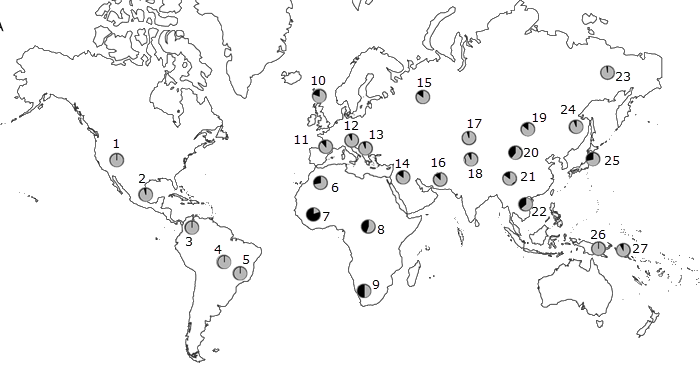
The data presented here are the allele frequencies for the five genes under consideration. The frequencies reflect the occurrence of the variant associated with lighter skin. These data are represented by maps in the slide set.
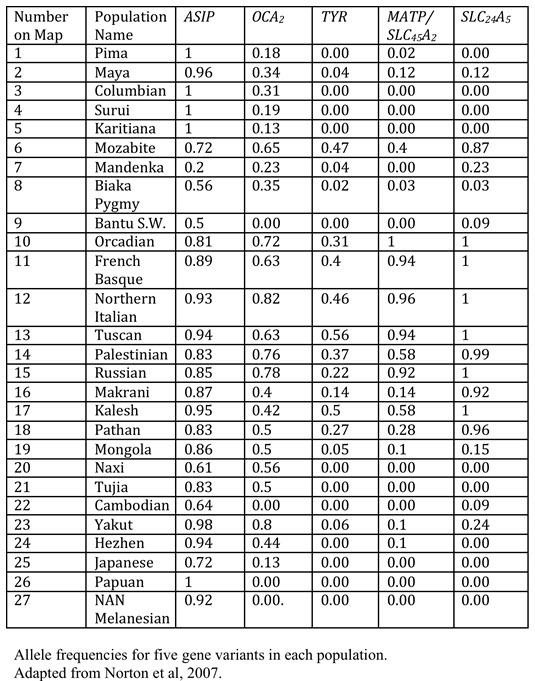
Summary of Norton data:
- ASIP gene: The variant associated with lighter skin is common among populations across the Northern hemisphere and is found in New World populations as well. The variant associated with darker skin is found in African and a few Asian populations.
- TYR gene: The variant associated with lighter skin is found predominantly in European populations and is absent in Asian populations with lighter skin.
- OCA2 gene: A version of this gene is associated with lighter skin in Northern hemisphere populations in Europe and across Asia.
- MATP = SLC45A2 gene: The variant associated with lighter skin is characteristic of western European populations, at a very high percentage.
- SLC24A5gene: The variant associated with lighter skin is found throughout Europe and the Near East.
What do these data indicate?
Populations with lighter skin commonly share variants for two genes, ASIP and OCA2. These are important for global patterns in skin color.
The SLC24A5,andSLC45A2variants are common in Europeans, but not East Asians.
TYR is stable around the world, except for western Europe. The lightest skin color (eg. Northern England) is associated with the lighter versions of all genes under consideration here.
Are Races Biological of Societal?
 As applied to human populations, race is a societal construct, not a biological concept. Human brains are good at putting things into categories. Two are Us and Them, which leads to separation along cultural dichotomies such as sports teams, clothing, customs and “race”. Racial concepts are not innate, but emerge from cultural biases.
As applied to human populations, race is a societal construct, not a biological concept. Human brains are good at putting things into categories. Two are Us and Them, which leads to separation along cultural dichotomies such as sports teams, clothing, customs and “race”. Racial concepts are not innate, but emerge from cultural biases.
Are There Races Among Humans?
Some biologists, primarily entomologists and plant biologists, apply the term “race” to populations that vary significantly from one another. These variations include chromosomal, genetic, geographical and/or physiological differences. Most vertebrate biologist use the term “subspecies” to denote differences within a species, rather than “races”.
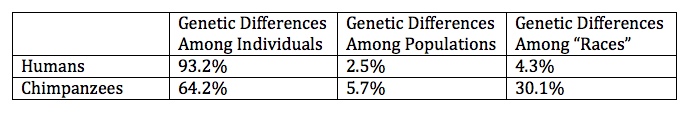
Subspecies have 1) sharp genetic boundaries among them and/or 2) separate evolution lineages, as evidenced by phylogenetic and genetic analysis.
The genetic differences within individual humans in a population (93.2%) are much greater than the genetic differences among populations (2.5%) and greater than the genetic differences among geographically defined “races” (4.3%). Comparatively, chimpanzees have less genetic variation among individuals in a population (64.2%) and more genetic differences among geographically delineated “races” (30.1%). These data show that there are no sharp genetic boundaries (=genetic differences) among human populations and that chimpanzees are much more genetically diverse across geographical boundaries.
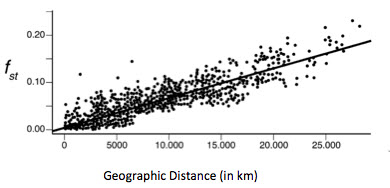
Analysis of population genetic data shows that the second attribute of subspecies, having separate evolutionary lineages, does not apply to humans. If this were true, graphing data related to differences in DNA would produce a graph with discontinuities or “breaks” in geographical distributions of humans (see graph below). Comparing genetic differences among and between human populations (known as the FST statistic) shows the opposite: continuous, smooth and relatively small differences. These data reflect the tendency of humans to migrate and interbreed, resulting in the mixing of genetic information.
But humans nonetheless categorize one another (and ourselves) by “race”, which itself is often correlated to skin color. Since genetic data do not support the idea of human races (or “subspecies”), race is a social construct, persistent in societies, both ancient and modern. If “race” is indeed a construct that is unsupported by a genetic analyses among humans, why are we prone to making these kinds of categorizations?
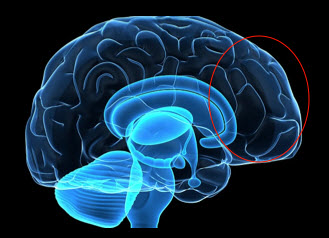 The human brain instantaneously categorizes things, including surface features of humans. This response leads to categorizing others as Us and Them. Categorizing occurs within milliseconds and begins with the neural input to the amygdala. The amygdala mediates unconscious emotions such as aggression, fear and anxiety. These are responses to something “different” in the environment and are not always accurate and often inappropriate. Information from the amygdala is forwarded to the prefrontal cortex, which mediates responses to social situations. The prefrontal cortex sorts things out – follows rules and puts things in categories- when presented with information from the amygdala, other neural inputs, and hormones. It integrates emotional and cognitive aspects of social life. In terms of race, this processing is highly variable and has a very strong cultural bias.
The human brain instantaneously categorizes things, including surface features of humans. This response leads to categorizing others as Us and Them. Categorizing occurs within milliseconds and begins with the neural input to the amygdala. The amygdala mediates unconscious emotions such as aggression, fear and anxiety. These are responses to something “different” in the environment and are not always accurate and often inappropriate. Information from the amygdala is forwarded to the prefrontal cortex, which mediates responses to social situations. The prefrontal cortex sorts things out – follows rules and puts things in categories- when presented with information from the amygdala, other neural inputs, and hormones. It integrates emotional and cognitive aspects of social life. In terms of race, this processing is highly variable and has a very strong cultural bias.
“Us” and “Them” categories are fluid and changeable; this particularly applies to racial concepts. Racial concepts are not innate, but emerge from cultural biases. These biases overlay brain processing that generically identifies “Us” and “Them”. Thus, human “race” is an idea, a concept, steeped in societal and historical norms. It is variable, fluid and ultimately context dependent.

Figure (above):These two young women, Lucy and Maria, were born at the same time to the same mother and father. They have other siblings, each with distinctive skin color and hair texture.
Lucy identifies herself as white and Maria identifies herself as black. They are using cultural definitions that are not based on a quantitative genetic analysis to define their identities.
The concept of human races is a powerful idea. It is the basis for racial profiling, racial identity, racial prejudice, racial pride, racial politics etc. The race concept can be a basis for both cohesion (Us) and divisiveness (Them). Humans identify others and themselves based on perceived ideas of race. Historical circumstances contribute to the definition of “race”. Since these circumstances change over time, so has the notion of “race”, as reflected in various attempts to collect census data. The egregious use of racial concepts, racism, is responsible for some of the worst behavior exhibited by humans: slavery and genocide.
References:
- Kubota, et al. 2012. The Neuroscience of Race. Nature Neuroscience 15(7): 940 – 946.
- Alan R. Templeton. 2013. Biological races in humans. Studies in History and Philosophy of Biological and Biomedical Sciences 44: 262-271.
- Robert M. Sopolsky. (2017). Behave: the biology of humans at our best and worst. Penguin Press.