
Monkey Opsins
 Humans have trichromatic vision. Our eyes detect colors in the visible range of the spectrum using three different cone cell types, each maximally stimulated by a different wavelength of light. Some humans are colorblind, meaning that their eyes detect colors in a more limited region of the spectrum using only one or two different types of cone cell.
Humans have trichromatic vision. Our eyes detect colors in the visible range of the spectrum using three different cone cell types, each maximally stimulated by a different wavelength of light. Some humans are colorblind, meaning that their eyes detect colors in a more limited region of the spectrum using only one or two different types of cone cell.
Color vision capabilities in the world’s two major groups of monkey species are patterned in the same way. Generally, monkey species from Africa, Europe and Asia (the old world monkeys) have trichromatic vision, whereas monkey found in South and Central America (the new world monkeys) have dichromatic vision.
This case will examine the evolution of trichromatic vision in old world monkeys.
Ecology: The Food Selection Hypothesis
Do trichromats gain a selective advantage through increased foraging efficiency?
If so, then we would predict that they would perform better in color-related tasks than their dichromatic counterparts. Conversely, in tasks where color discrimination is not important, we would predict that dichromats would perform equally as well, or better, than trichromats.

To see whether or not color-based selection was occurring, Smith et al (2003) tested trichromat and dichromat foraging ability when presented ripe and unripe fruit hidden amongst green leaves. In their experiment, trichromatic monkeys selected the ripe fruit about 53% of the time whereas dichromatic monkeys did so about 37% of the time. This study supports the hypothesis that, in some situations, trichromatic vision gives an advantage for ripe-fruit foraging.
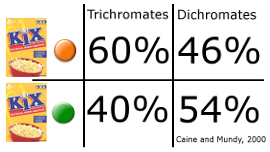 In another experiment, Caine and Mundy (2000) had trichromat and dichromat monkeys forage for colored pieces of cereal scattered and hidden in different colors of shrubbery and grass. They found that trichromats found significantly more orange pieces of cereal in a green environment where color distinction was difficult for dichromats. However, they also found that dichromats found more green pieces of cereal in the green environment. Although trichromats had an advantage in finding food in some cases, the effect was somewhat dependent on the environment. When the color of the food matched the surroundings, dichromats actually performed better than trichromats.
In another experiment, Caine and Mundy (2000) had trichromat and dichromat monkeys forage for colored pieces of cereal scattered and hidden in different colors of shrubbery and grass. They found that trichromats found significantly more orange pieces of cereal in a green environment where color distinction was difficult for dichromats. However, they also found that dichromats found more green pieces of cereal in the green environment. Although trichromats had an advantage in finding food in some cases, the effect was somewhat dependent on the environment. When the color of the food matched the surroundings, dichromats actually performed better than trichromats.
Some experiments suggest that there are times when trichromatic vision may be disadvantageous to foraging efficiency. In a study by Saito et al (2005) dichromat monkeys were significantly more efficient than trichromat monkeys in picking out shapes that were camouflaged in red and green to match a red and green background. Trichromat monkeys were distracted by the contrasting colors in the camouflage pattern, but to dichromats the camouflage looked like one single color. This apparently allowed the dichromats to pick out the shapes more easily.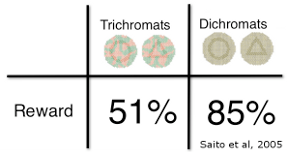
Conclusion
The evidence seems to indicate that trichromat monkeys have a foraging advantage when it comes to choosing ripe fruit over unripe fruit. They also seem more able to pick out non-green fruit amongst green leaves. Dichromat monkeys seem to have a foraging advantage when it comes to choosing green food amongst green leaves. They also tend to have more rod cells than trichromat monkeys that may make them more efficient foragers in low light.
Cell Biology
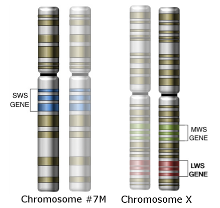
The spectrum of colors a monkey can detect depends on how many kinds of light sensitive opsin proteins that the cone cells in its eye are able to produce. Each cone cell contains only one kind of opsin. Dichromats have two opsin genes and can make two opsin proteins – the short wave sensitive opsin protein (SWS) and the medium wave sensitive opsin protein (MWS). Trichromats have three opsin genes, one coding for an additional opsin protein – the long wave sensitive opsin protein (LWS).
Process Overview
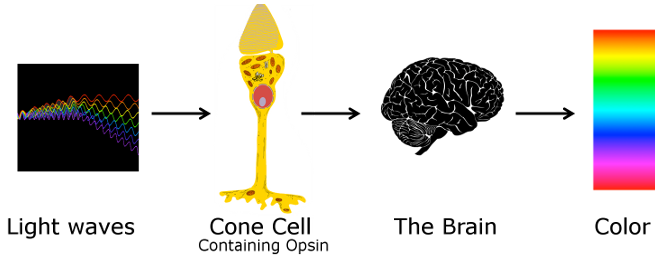
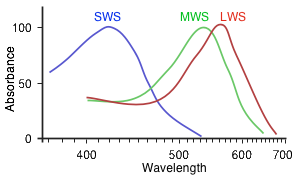
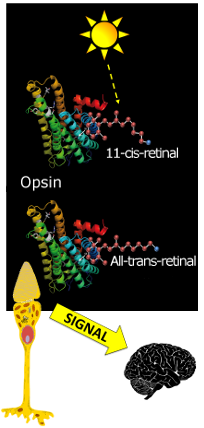 An opsin protein is a transmembrane protein in a cone cell. It is stimulated when a photon of light enters the eye and activates a chromophore molecule, a retinal, embedded in the opsin. When the retinal chromophore is activated it changes shape (configuration) from an 11-cis arrangement to an all-trans arrangement. Once the chromophore is in an all-trans state, the opsin protein it is associated with is stimulated and the cone cell emits a signal that ultimately travels to the brain indicating that light has been detected. Because the three different types of opsin (SWS, MWS and LWS) are structurally different, they each interact with the retinal chromophore in a slightly different way. This results in chromophore activation and opsin stimulation at different wavelengths for each different type of opsin. For SWS opsin proteins, retinal chromophores are most efficiently activated when light entering the eye has a wavelength of 420 nm (blue; see the spectrum below). Retinal chromophores in SWS opsin proteins can be activated for light as short as ~360 nm and as long as ~540 nm but retinal activation is most efficient (i.e. it peaks) at 420 nm. For MWS opsin proteins, retinal chromophores can be activated by light with wavelengths between ~400 nm and 650 nm, peaking at 534 nm (green). For LWS opsin proteins, retinal chromophores can be activated by light with wavelengths between 400 nm and 680 nm, peaking at 564 nm (darker green). The longer wavelengths allow the observer to perceive red.
An opsin protein is a transmembrane protein in a cone cell. It is stimulated when a photon of light enters the eye and activates a chromophore molecule, a retinal, embedded in the opsin. When the retinal chromophore is activated it changes shape (configuration) from an 11-cis arrangement to an all-trans arrangement. Once the chromophore is in an all-trans state, the opsin protein it is associated with is stimulated and the cone cell emits a signal that ultimately travels to the brain indicating that light has been detected. Because the three different types of opsin (SWS, MWS and LWS) are structurally different, they each interact with the retinal chromophore in a slightly different way. This results in chromophore activation and opsin stimulation at different wavelengths for each different type of opsin. For SWS opsin proteins, retinal chromophores are most efficiently activated when light entering the eye has a wavelength of 420 nm (blue; see the spectrum below). Retinal chromophores in SWS opsin proteins can be activated for light as short as ~360 nm and as long as ~540 nm but retinal activation is most efficient (i.e. it peaks) at 420 nm. For MWS opsin proteins, retinal chromophores can be activated by light with wavelengths between ~400 nm and 650 nm, peaking at 534 nm (green). For LWS opsin proteins, retinal chromophores can be activated by light with wavelengths between 400 nm and 680 nm, peaking at 564 nm (darker green). The longer wavelengths allow the observer to perceive red.
The Opsin Protein
The opsin protein is composed of a string of amino acids. Each green dot in the 2D visualization represents one amino acid. The LWS opsin functionally differs from the MWS opsin in only three places in the amino acid sequence. When amino acid # 180 is an alanine, amino acid # 277 is a phenylalanine and amino acid # 285 is an alanine the resulting opsin protein has an stimulation maximum of ~ 534 nm (i.e. it is medium wave sensitive, MWS). When amino acid # 180 is a serine, amino acid # 277 is a tyrosine and amino acid # 285 is a threonine the resulting opsin protein has a stimulation maximum of ~ 564 nm (i.e. it is long wave sensitive, LWS).
 The responses to light of each opsin protein (SWS, MWS and LWS) in trichromats are shown to the right. Note how similar the curves look for MWS and LWS. The LWS stimulation curve is shifted 30 nm to the right of the MWS stimulation curve. Dichromats, who lack the LWS, are not able to “see” colors of longer wavelength, and thus miss out on some of the redder hues.
The responses to light of each opsin protein (SWS, MWS and LWS) in trichromats are shown to the right. Note how similar the curves look for MWS and LWS. The LWS stimulation curve is shifted 30 nm to the right of the MWS stimulation curve. Dichromats, who lack the LWS, are not able to “see” colors of longer wavelength, and thus miss out on some of the redder hues.
Molecular Genetics
 Humans have trichromatic vision. This means that our eyes can detect color at three different color wavelengths: blue, green and red. This is not true of all primates. Some have dichromatic vision, they can only detect two different wavelengths.
Humans have trichromatic vision. This means that our eyes can detect color at three different color wavelengths: blue, green and red. This is not true of all primates. Some have dichromatic vision, they can only detect two different wavelengths.
DNA, Central Dogma and the Opsin Protein
DNA is made up of long strings of nucleotides. There are four nucleotides in DNA: Adenine (A), Thymine (T), Guanine (G) and Cytosine (C). These nucleotides are held together by a phosphate-sugar “backbone” and are complemented by a parallel strand of nucleotides making a DNA molecule “double-stranded”. Parallel strands are held together by hydrogen bonds between bases; Adenine binds to Thymine, Cytosine binds to Guanine. Discrete sections of DNA carry sequences of nucleotides that, when processed, are responsible for proteins or functional RNA chains. These sections are called genes. When making a protein, the DNA is first transcribed into a single-stranded piece of messenger RNA (mRNA) using an enzyme called RNA polymerase. The newly made mRNA is then translated through a ribosome into a long chain of amino acids. This chain of amino acids folds up to make a functional protein. Proteins build the organism, including all measurable and observable characteristics.
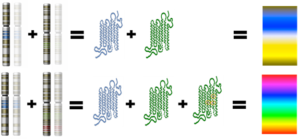
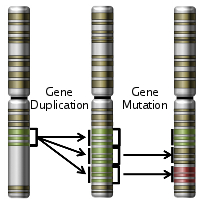 In the case of monkeys, the SWS opsin gene is located on an autosome (non sex chromosome) – chromosome #7. The MWS and LWS opsin genes are located on the X-chromosome. In many primate species there are many non-functional copies of the MWS gene found on the X-chromosome. This indicates that there were duplication events of the MWS gene that occurred many times throughout the history of trichromat monkey species. One of those MWS copies accumulated a few specific DNA nucleotide substitution mutations, which gave rise to an LWS opsin gene that encoded a functional LWS opsin protein. In essence, only three specific mutations were needed over tens of millions of years in order for a functional LWS gene to successfully evolve.
In the case of monkeys, the SWS opsin gene is located on an autosome (non sex chromosome) – chromosome #7. The MWS and LWS opsin genes are located on the X-chromosome. In many primate species there are many non-functional copies of the MWS gene found on the X-chromosome. This indicates that there were duplication events of the MWS gene that occurred many times throughout the history of trichromat monkey species. One of those MWS copies accumulated a few specific DNA nucleotide substitution mutations, which gave rise to an LWS opsin gene that encoded a functional LWS opsin protein. In essence, only three specific mutations were needed over tens of millions of years in order for a functional LWS gene to successfully evolve.
 Sex Linked Trait
Sex Linked Trait
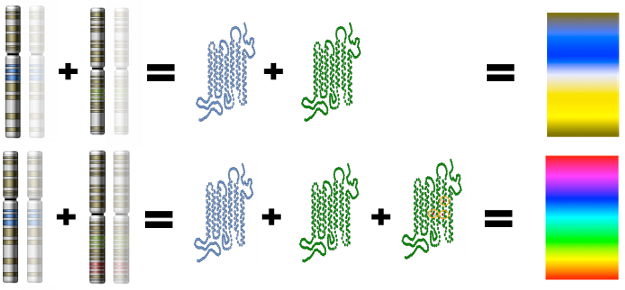
Because the MWS and LWS genes are on the X chromosome, females have two copies of each whereas males have only one of each. New world monkeys usually only have the MWS gene, lacking the L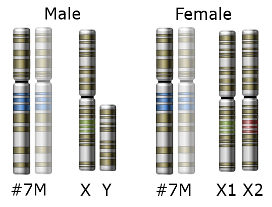 WS gene. Nonetheless, some new world monkey species have some trichromatic females and only dichromatic males. This occurs when there are different variants (i.e. alleles) of the MWS gene, one on each X chromosome, that are spectrally tuned to different wavelengths. Females that have two MWs alleles are therefore trichromatic. Females that have only one allele are dichromatic. Males can only be dichromatic because they only have one x-chromosome.
WS gene. Nonetheless, some new world monkey species have some trichromatic females and only dichromatic males. This occurs when there are different variants (i.e. alleles) of the MWS gene, one on each X chromosome, that are spectrally tuned to different wavelengths. Females that have two MWs alleles are therefore trichromatic. Females that have only one allele are dichromatic. Males can only be dichromatic because they only have one x-chromosome.
Heterozygous females of these species have three different opsin genes that code for three different opsin proteins, one from each X chromosome and one from chromosome 7. Homozygous females and all males of these species have only two variations of on the opsin gene so they express only two opsin proteins, one from the X chromosome and one from chromosome 7 (see Jacobs, 1996 for more on trichromatic females and see Neitz and Neitz, 2000 for more on spectral tuning of opsin proteins).
Phylogenetics
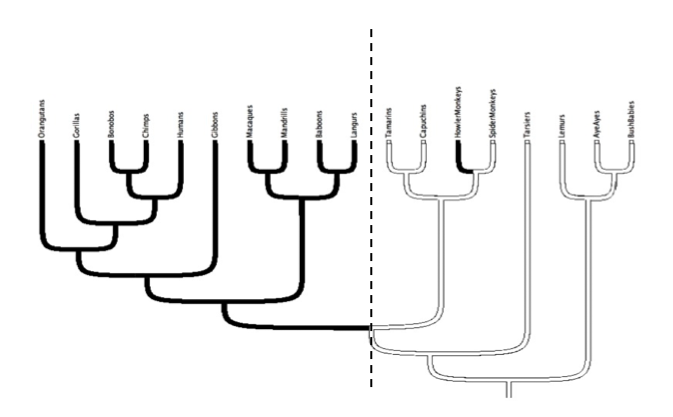 A phylogenetic tree is a way to represent the genetic relatedness among species in an evolutionary context (see Baum et al 2005). When we construct a phylogenetic tree for monkeys, based on morphological, behavioral, or DNA sequence characteristics, we find that the old world monkey species (trichromats) form a cohesive monophyletic group (or clade) on one side of the tree while the new world monkey species (dichromats) form a clade on the other side of the tree. The presence of these two clades indicates that the new world monkeys separated from the old world monkeys at some point in their history. Our observation that old world monkeys have trichromatic vision and new world monkeys have dichromatic vision implies that trichromatic vision evolved in the old world monkeys after this separation event.
A phylogenetic tree is a way to represent the genetic relatedness among species in an evolutionary context (see Baum et al 2005). When we construct a phylogenetic tree for monkeys, based on morphological, behavioral, or DNA sequence characteristics, we find that the old world monkey species (trichromats) form a cohesive monophyletic group (or clade) on one side of the tree while the new world monkey species (dichromats) form a clade on the other side of the tree. The presence of these two clades indicates that the new world monkeys separated from the old world monkeys at some point in their history. Our observation that old world monkeys have trichromatic vision and new world monkeys have dichromatic vision implies that trichromatic vision evolved in the old world monkeys after this separation event.
Given what we know about geology, paleontology and ancient primate history we can start to map other events onto the phylogenetic tree.
- The first primate species evolved sometime between 70 and 80 million years ago (e.g. Chatterjee et al 2009).
- There were primate species that lived throughout the contiguous new world and old world landmasses around 55 million years ago (see: Oldest North American Primate Fossil Discovered).
- The new and old world landmasses separated sometime around 50 million years ago.
- The successful gene duplication and mutation events of the MWS opsin that resulted in a function LWS opsin happened sometime around 40 million years ago in an ancient old world primate lineage Surridge et al 2003).
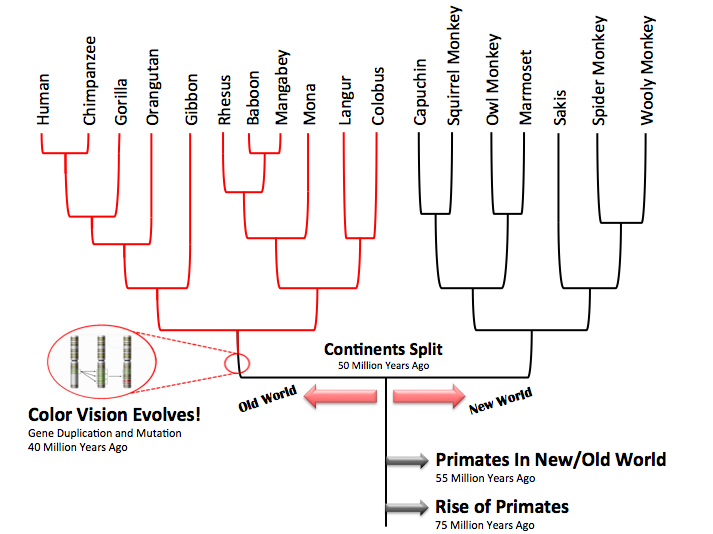 You may notice that the above phylogenetic tree does not include all primate species but is instead a representative tree of three different primate groups. The dichromate phylogenetic group (i.e. cluster of branches) the right shows the new world monkeys (platyrrhines). The trichromatic phylogenetic groups to the left show the apes (hominoids) and old world monkeys (cercopithecids). While this case study has focused on the old world monkeys, the evolution of trichromatic vision actually occured before old world monkeys and apes were phylogenetically distinctive groups.
You may notice that the above phylogenetic tree does not include all primate species but is instead a representative tree of three different primate groups. The dichromate phylogenetic group (i.e. cluster of branches) the right shows the new world monkeys (platyrrhines). The trichromatic phylogenetic groups to the left show the apes (hominoids) and old world monkeys (cercopithecids). While this case study has focused on the old world monkeys, the evolution of trichromatic vision actually occured before old world monkeys and apes were phylogenetically distinctive groups.